
Page 124 - 《中国药房》2023年1期
P. 124
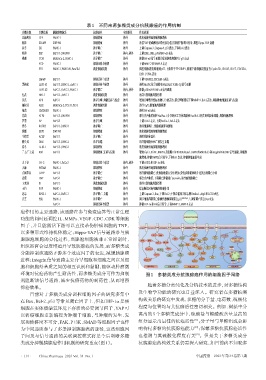
表1 不同来源多糖类成分抗胰腺癌的作用机制
多糖来源 多糖名称 胰腺癌细胞系 抗癌途径 实验阶段 作用机制
北极海参 CFP PANC-1 抑制增殖 体外 浓度依赖性地抑制细胞增殖
刺参 SJAMP SW1990 抑制增殖 体外 改善YAP 的磷酸化和染色质易位并抑制下游基因转录,调控Hippo-YAP 通路
茯苓 EE PANC-1 诱导凋亡 体外 上调Caspase-3、Caspase-9、p53表达,下调Bcl-2表达
海胆 SEP BxPC-3、SW1990 诱导凋亡 体内、体外 上调ERK、JNK、p38和NF-κB表达
褐藻 TCF5 MIAPaCa-2、PANC-1 诱导凋亡 体外 抑制NF-κB信号通路并促进肿瘤抑制因子p53表达
FCD PANC-1 抑制迁移与侵袭 体外 下调MMP-2 和 MMP-9表达
FV1 PANC-1、PANC-89、PancTu1 调控细胞周期 体外 调控细胞周期抑制剂p57、上游因子TP53INP1,抑制下游细胞周期蛋白Cyclin E2、CDC45、CDC7、CDC25A、
E2F1、PCNA表达
LMWF BxPC-3 抑制迁移与侵袭 体外 下调VEGFR2、PECAM-1表达
黑枸杞 LRP3-S1 BxPC-3、PANC-1、AsPC-1 抑制迁移与侵袭 体外 调控MAPK信号通路和FAK/AKT /GSK-3β信号通路
LRP1-S2 AsPC-1、BxPC-3、PANC-1 诱导凋亡 体内、体外 抑制 p38 MAPK/NF-κB信号通路
红花 HH1-1 BxPC-3、AsPC-1 调控细胞周期 体外 诱导S期细胞周期停滞
黄芪 APS AsPC-1 诱导自噬、抑制迁移与侵袭 体外 增加自噬特异性标志物LC3的表达;联合阿帕替尼下调MMP-9、Bcl-2表达,抑制肿瘤细胞迁移与侵袭
姬松茸 ABE MIAPaCa-2、PCI-35、PK-8 调控细胞周期 体外 诱导G 0/G 1期细胞周期阻滞
菊花 CMTA0S3 PANC-1 抑制增殖 体外 调控NF-κB表达
亮菌 ATPS BxPC-3、SW1990 抑制增殖 体外 联合吉西他滨调控Fas/Fas-L介导的死亡受体通路和Bax/Bcl-2相关的线粒体通路,抑制细胞增殖
芦荟 AP BxPC-3 诱导自噬 体外 上调ULK1表达,下调Beclin-1、Bcl-2表达
麦冬 ROH05 BxPC-3、PANC-1 诱导凋亡 体外 诱导细胞凋亡、细胞核破碎和皱缩
泥螺 BEP3 SW1990 抑制增殖 体外 浓度依赖性地抑制细胞增殖
牛樟芝 ACEP BxPC-3 诱导凋亡 体外 调控线粒体途径
蒲公英 DRE BxPC-3、PANC-1 诱导自噬 体外 诱导癌细胞在死亡前发生自噬
忍冬 LJ-02-1 BxPC-3、PANC-1 抑制增殖 体外 浓度依赖性地抑制细胞增殖
三七/三七花 RN1 BxPC-3 抑制增殖、迁移与侵袭 体外 靶向Gal-3、EGFR、BMPRs并阻断EGFR/ERK/Runx1、BMPR/SMAD/Id-3和integrin/FAK/JNK信号通路,抑制细
胞增殖;抑制BMP2信号转导,下调Id-1表达,抑制肿瘤血管生成
太子参 H-1-2 PANC-1、PaCa-2 抑制迁移与侵袭 体内、体外 下调AGR2和HIF-1α表达
天麻 WTMA PANC-1 抑制增殖 体外 浓度依赖性地抑制细胞增殖
喜树果实 CAFP BxPC-3 诱导凋亡 体外 诱导癌细胞凋亡,使细胞收缩百分比增加、染色质浓缩和碎片化及出现凋亡小体
香菇 LNT AsPC-1 诱导凋亡 体外 联合吉西他滨,下调凋亡抑制蛋白survivin,诱导癌细胞凋亡
玉米须 S1 BxPC-3 调控细胞周期 体外 诱导S期细胞周期停滞
玉竹 POP PANC-1 抑制增殖 体外 提高顺铂对癌细胞的抑制作用
远志 RP02-1 AsPC-1、BxPC-3 诱导凋亡、自噬 体外 上调Caspase-3、Bax,下调Bcl-2;干扰自噬相关标志物Beclin-1、Atg5和 LC3B表达
云芝 PSK PANC-1 诱导凋亡 体外 调节细胞周期蛋白依赖性激酶抑制蛋白p21 WAF/Cip1 ,上调促凋亡蛋白Bax表达
AsPC-1 抑制迁移与侵袭 体外 抑制HIF-1α和Hh 信号转导,下调MMP-2、MMP-9表达
癌作用的主要通路,该通路在参与免疫应答等正常生理
功能的同时还调控IL、MMPs、VEGF、CDC、CDK等细胞
因子,并且能激活下游可以直接杀伤肿瘤细胞的 TNF,
以多种形式维持机体稳定;Hippo-YAP信号通路参与胰
腺腺泡细胞的分化过程,当腺泡细胞被非正常抑制时,
组织器官会过度增殖并导致胰腺癌的发展,而多糖类成
分能抑制该通路下游多个效应因子的表达,减缓胰腺癌
进程;Integrin信号通路主要介导细胞和细胞之间以及细
胞和细胞外基质之间的相互识别和黏附,能驱动肿瘤微
环境对抗癌药物产生耐药性,而多糖类成分可作为抑制 图1 多糖类成分抗胰腺癌作用的细胞因子网络
剂阻断该信号通路,减少抗癌药物的耐药性,从而增强
随着多糖分离纯化及分析技术的进步,对多糖结构
抑癌效果。
目前对于多糖类成分调控细胞因子的研究多集中 及生物学功能的研究也日益深入。研究者在多糖抗癌
在Bax、Bcl-2、p53等常见凋亡因子上,但如HIF-1α 是癌 构效关系的研究中发现,多糖的分子量、电荷数、硫酸化
细胞在实体瘤缺氧环境下存活的必要调节因子,YAP可 程度与位置均与其抗癌活性密切相关。例如:褐藻中分
以将癌细胞重新编程为肿瘤干细胞,与肿瘤的发生、发 离出的5个多糖类成分中,硫酸盐与糖醛酸含量最高的
[15]
展和转移密不可分,FAK、P13K、SMAD等细胞因子也作 组分显示出最佳的抗癌活性 ;分子量与单糖组成会影
[39]
为中间递质参与了多条抑制胰腺癌的途径,这些细胞因 响枸杞多糖的抗胰腺癌能力 ;泥螺多糖抗胰腺癌活性
[27]
子间及与信号通路的关联调控模式将是今后明晰多糖 也推测与其硫酸化程度有关 。但是关于多糖类成分
类成分抑制胰腺癌作用机制的研究重点(图1)。 抗胰腺癌的构效关系仍需深入研究,如目前尚不知晓多
· 114 · China Pharmacy 2023 Vol. 34 No. 1 中国药房 2023年第34卷第1期