
Page 133 - 《中国药房》2021年07期
P. 133
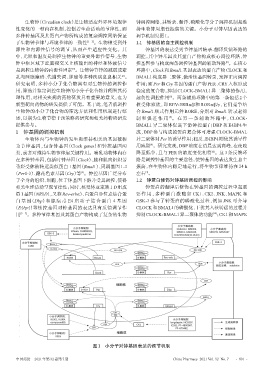
生物钟(Circadian clock)是生物适应外界环境规律 钟调控网络,其转录、翻译、磷酸化等分子调控机制是维
性变化的一种内在机制,控制着生命活动的节律性,而 持生物钟周期性振荡的关键。小分子对钟基因表达的
多种钟基因及其蛋白产物所构成的复杂调控网络保证 调节机制见图1。
了生物钟节律与环境节律的一致性 [1-2] 。生物钟受到外 1.1 钟基因的自主调控机制
源性和内源性信号的调节,从而产生适应性变化。其 钟基因的表达受到节律基因转录-翻译反馈环路的
中,光照和温度是调控生物钟的重要外源性信号,生物 调控,其中钟基因及其蛋白产物构成核心调控环路,钟
钟中枢区域下丘脑视交叉上核输出的神经和体液信号 控基因参与构成两条调控钟基因的辅助环路 。在核心
[8]
[2]
是调控生物钟的内源性因素 。生物钟节律的减弱或紊 环路中,Clock和Bmal1基因表达的蛋白产物CLOCK和
[3]
乱与睡眠障碍、代谢失调、肿瘤等多种疾病息息相关 。 BMAL1构成异二聚体,激活钟基因转录,发挥正向调控
研究表明,多种小分子化合物具有对生物钟的调控作 作用;而Per和Cry基因的蛋白产物PER、CRY入核形成
用,筛选并鉴定调控生物钟的小分子化合物并阐明其药 稳定的复合物,抑制 CLOCK-BMAL1 异二聚体的作用,
理作用,对相关疾病的药物研发具有重要的意义,也为 起负性调控作用 。两条辅助环路中的第一条包括2个
[8]
新型靶向药物的研发提供了可能。基于此,笔者就调控 核受体家族,即 REV-ERBα/β和 RORα/β/γ,它们竞争结
生物钟的小分子化合物的筛选方法和作用机制进行综 合 Bmal1 顺式作用元件 RORE,分别对 Bmal1 转录起抑
述,以期为生物节律干预策略的研究和相关药物的研发 制 和 促 进 作 用 。 在 另 一 条 辅 助 环 路 中 ,CLOCK-
[5]
提供参考。 BMAL1 异二聚体促进下游钟控蛋白 DBP 和 E4BP4 生
1 钟基因的调控机制 成,DBP 参与构成的蛋白复合体可增强 CLOCK-BMAL
生物体内与生物钟的发生和维持相关的基因被称 异二聚体对Per的诱导作用;相反,E4BP4则使其诱导作
[9]
为节律基因,包含钟基因(Clock genes)和钟控基因两 用减弱 。研究发现,DBP浓度在清晨达到高峰,在夜晚
[4]
类,前者对维持生物节律起关键作用。哺乳动物体内存 降至低谷,且与 PER 的浓度变化相符 。这 3 条反馈环
在多种钟基因,包括时钟基因(Clock)、脑和肌肉组织芳 路是调控钟基因的主要途径,使钟基因的表达发生自主
香羟受体核转运类似蛋白1基因(Bmal1)、周期基因1-3 振荡,在生物体内稳定地运行,将生物节律维持在 24 h
[4]
[4]
(Per1-3)、隐花色素基因(Cry)等 。钟控基因广泛分布 左右 。
于全身的组织、细胞,位于钟基因下游并受其调控,使得 1.2 钟蛋白修饰对钟基因调控的影响
相关生理活动呈现节律性;同时,核受体亚家族1 D组成 钟蛋白的翻译后修饰在钟基因的调控过程中起重
员1基因(NR1D1,又称Rev-erbα)、白蛋白D位点结合蛋 要作用,多种蛋白激酶如 CK1、CK2、JNK、MAPK 和
白 基 因(Dbp)和 腺 病 毒 E4 启 动 子 结 合 蛋 白 4 基 因 GSK-3 参与了钟蛋白的磷酸化过程,例如 JNK 可介导
(E4bp4)等钟控基因对钟基因的表达具有反馈调节作 CLOCK和BMAL1的磷酸化、干扰其入核转运的过程并
[10]
用 [5-7] 。多种节律基因及其蛋白产物构成了复杂的生物 抑制CLOCK-BMAL1异二聚体的功能 ;CK1和MAPK
图1 小分子对钟基因表达的调节机制
中国药房 2021年第32卷第7期 China Pharmacy 2021 Vol. 32 No. 7 ·891 ·